农杆菌介法将耐盐基因HAL1转入玉米自交系H99的研究 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 22 篇 doi: 10.5376/mpb.cn.2011.09.0022
收稿日期: 2010年12月22日 接受日期: 2011年02月18日 发表日期: 2011年02月28日
孙传波等, 2011, 农杆菌介法将耐盐基因HAL1转入玉米自交系H99的研究, 分子植物育种 Vol.9 No.22 (doi: 10.5376/mpb.cn.2011.09.0022)
通过农杆菌介导法将耐盐基因HAL1转入玉米自交系H99中,同时研究了愈伤组织状态、农杆菌浓度、侵染时间和真空压力等因素对转化频率的影响。结果表明:继代5 d的愈伤组织、菌液浓度OD600为0.6和0.05 Mpa真空压力侵染时间15 min为最佳转化条件。通过该优化体系获得的转基因植株18株,经PCR分析鉴定,其中9株表现阳性,初步证明外源基因已经整合到玉米基因组中。
玉米(Zea mays L.)是三大粮食作物之一,在我国农业生产中占有重要地位,转基因玉米的研究也因此受到普遍重视。自从首次获得玉米转基因完整植株以来(Rhodes., 1988),玉米遗传转化技术得到了较大的发展,利用现代生物技术进行玉米品种改良的研究已日趋深入。目前玉米的遗传转化方法主要有基因枪法和农杆菌转化法 (袁英等, 2006),农杆菌作为一种天然的植物基因转化系统,具有转化的外源DNA结构完整、转化机理清楚、整合位点较稳定、拷贝数低、遗传稳定性好等优点。在国外通过农杆菌转化法获得了很多稳定的转基因玉米(Gould et al., 1991; Ishida et al., 1996; Huang et al., 2004),在国内也建立了农杆菌介导的玉米遗传转化体系(黄璐等, 1999)。
干旱、盐碱等逆境条件严重影响玉米的生产,利用转基因技术来改善玉米的抗逆性一直是生物学家的重要研究内容。HAL1为细胞离子平衡调节因子,其过表达使细胞内K+浓度提高Na+浓度降低,提高细胞对盐的耐受性((Rios et al., 1997; Yang et al., 2001)。国外率先将HAL1基因导入甜瓜(Bordas et al., 1997),提高了转基因植株在组织培养条件下的耐盐性,随后HAL1基因在番茄、烟草和水稻等植物中也得到了表达(田吉林等, 2003; Rohila et al., 2002),耐盐检测表明,转HAL1基因植株的耐盐性比野生型有明显提高。这表明,通过HAL1基因改良生物的耐盐性是有效的。本文研究愈伤组织状态、农杆菌浓度、侵染时间及真空压力等因素对玉米农杆菌遗传转化的影响,建立适宜自交系H99的农杆菌转化体系,通过体系将耐盐基因HAL1转入玉米自交系H99,获得抗旱耐盐的优良玉米种质资源。
1结果与分析
1.1愈伤组织状态对遗传转化的影响
以继代不同时间的胚性愈伤组织为外植体进行农杆菌转化, 经3次筛选后,所得到的抗性愈伤组织率不同。由表1的实验结果可以发现,胚性愈伤组织状态明显的影响着抗性愈伤率,继代5 d后转化的胚性愈伤组织其抗性愈率最高为38.46%,而继代0 d和继代12 d的胚性愈伤组织转化后抗性愈伤率分别为5.61%和10.26%,明显低于继代5d的胚性愈伤组织,由此可见选择继代5 d后的胚性愈伤组织最适宜农杆菌转化。
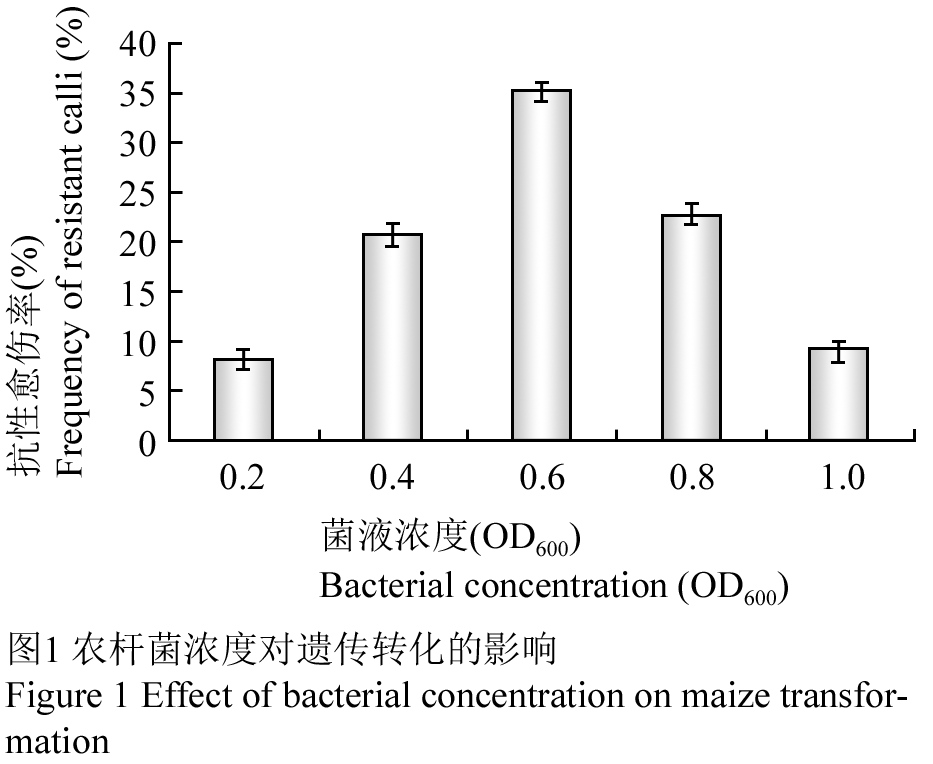
1.2农杆菌浓度(OD600)对遗传转化的影响
菌液浓度是影响遗传转化效率的关键因素之一,以继代5 d的愈伤组织为外植体,用不同OD600值的菌液转化胚性愈伤组织,经过3次抗性筛选后,统计抗性愈伤组织率。由图1结果可以明显发现,当OD600=0.6时抗性愈伤率最高为35.33%, OD600=0.2时最低位8.33%,当OD600小于0.6时随着菌液浓度增大,抗性愈伤率不断提高;当OD600大于0.6时随着菌液浓度增大,抗性愈伤率反而降低,这可能是由于高浓度菌体对愈伤组织的伤害大和共培养后难抑制菌造成的。
1.3真空压力和侵染时间对遗传转化的影响
|
Table 2 Effect of vacuum pressure and infection time on maize transformation
|
1. 4转基因植株的分子生物学检测
通过该优化体系转化1260块H99胚性愈伤组织,共获得转化植株18株,经过PCR初步鉴定,其中9株呈阳性,抗性植株阳性率为50%,图2结果显示,转基因植株扩增出375 bp的目的条带,和阳性对照一致,可以初步证明外源基因已经整合到玉米基因组中。
|
图2 转基因植株PCR分析 Figure 2 PCR analysis on transgenic plants 注: M: DL2000; -: 阴性对照; +: 阳性对照; 1-9: 转基因植株 Notes: M: DNA marker; -: Negative control; +: Positive control; 1-9: Transgenic plants |
2讨论
为了建立完善的农杆菌介导的玉米遗传转化体系,本实验以H99胚性愈伤组织为外植体,研究了胚性愈伤组织状态、农杆菌浓度、侵染时间和真空压力等因素对转化效率的影响,并应用该体系将耐盐基因HAL1转入自交系H99中。试验中发现,胚性愈伤组织状态对转化效率影响非常明显,当以继代5d的胚性愈伤组织为外植体,抗性愈伤获得率高于其他条件,这说明此状态的愈伤组织刚好处于对数生长期,比较利于转化。菌液浓度对转化频率有着直接影响,当OD600=0.6时抗性愈伤率最高为35.33%,但是当浓度过高时(OD600>0.6),由于吸附得菌液过多,对愈伤组织的伤害增大,后期的抑制菌难度增加,转化率反而降低。在本实验中还尝试了真空压力对遗传转化的影响,在0.05 Mpa和0.07Mpa真空压力条件下,侵染15 min和20 min的抗性愈伤率几乎相同,但是发现同等压力条件下,侵染时间长的难抑制菌,而且状态不好;相同侵染时间时,真空压力越大越难抑制菌;综合考虑选择0.05 Mpa真空压力,侵染15 min的转化条件。因此选择继代5 d的愈伤组织、菌液浓度OD600=0.6和0.05 Mpa真空压力侵染时间15 min为目前本实验室的玉米农杆菌转化的转化条件,要建立一套转化效率高的完善体系,还需要研究影响遗传转化效率的其他因素,我们将在以后的试验中继续优化。
通过该优化体系将耐盐基因HAL1转入玉米自交系H99的愈伤组织(1 260块)中,经过3 d共培养,4 d恢复培养,三次除草剂(Bar3.0mg/L)筛选后,获得转基因玉米材料18株,经过PCR初步鉴定,其中9株呈阳性,抗性植株阳性率为50%,初步证明外源基因HAL1已经整合到玉米基因组中。转HAL1植株的获得将为玉米抗旱耐盐的新品系玉米品种培育奠定良好的基础。
3材料与方法
3.1 材料
3.1.1 植物材料
玉米自交系H99。
3.1.2 植物表达载体
植物载体为pC3301-HAL1,含有目的基因HAL和筛选标记基因Bar(图3)。
|
|
3.1.3培养基
实验中所用到的培养基如下:YEP培养基:酵母提取物10g/L + 蛋白胨 10g/L + NaCl 5 g/L,pH=7.0;侵染培养基:1/2 MS大量,1/2 MS微量,1/2 MS维生素,100 mg/L肌醇,水解酪蛋白500 mg/L,2,4-D 1.5 mg/L AS100 mg/L+3.6%蔗糖+ 6.8%葡萄糖,pH=5.2;愈伤组织诱导培养基:MS大量,MS微量,MS维生素,100 mg/L肌醇,水解酪蛋白500 mg/L,L-脯氨酸500 mg/L,L-天门冬酰胺200 mg/L,泛酸钙0.5 mg/L,生物素0.05 mg/L,2,4-D 1.5 mg/L,蔗糖 4%(w/v) pH=6.0;继代培养基:MS大量,MS微量,MS维生素,100 mg/L肌醇,水解酪蛋白500 mg/L, 2.4-D 2.0mg/L,蔗糖3%(w/v) pH=6.0;共培养培养基:1/2 MS大量,1/2 MS微量,1/2 MS维生素,2,4-D 1.5 mg/L AS100 mg/L AgNO3 850 mg/L+半光氨酸100 mg/L+3%蔗糖pH=6.0;恢复培养基:1/2 MS大量,1/2 MS微量,1/2 MS维生素,2,4-D1.5 mg/L +MES 0.5g+AS100 mg/L+Cef 500 mg/L+3%蔗糖pH=6.0;筛选培养基: 1/2 MSZ+2,4-D 1.5 mg/L +MES 0.5 g +Cef 500 mg/L+除草剂(Bar 3.0 mg/L)+3%蔗糖pH=6.0;分化培养基:继代培养基+KT 0.5 mg/L +Cef 500 mg/L代培养基+KT 0.5 mg/L +Cef 500 mg/L pH=5.8;生根培养基:继代培养基+IBA 0.5 mg/L +Cef 500 mg/L+活性炭5 g/L pH=6.0。
3.2方法
3.2.1 外植体的获取
取授粉9~13d,1mm左右的幼胚接种在诱导培养基上,每14d继代一次,继代3次后得到胚性愈伤组织。
3.2.2农杆菌转化
3.2.2.1工程菌的制备
将含有质粒pC3301-HAL1的农杆菌EHA105在含相应抗生素的YEP液体培养基中28℃, 180 rpm培养至OD600值在0.8左右,离心5 min收集菌体,然后用侵染培养基重悬菌体,加入乙酰丁香酮(AS) 100 μmol/L,调整重悬液浓度至OD600=0.6。
3.2.2.2 外植体的侵染
将继代0~12d的愈伤组织分别集中于小三角瓶中,用OD值为0.2、0.4、0.6、0.8、1.0的重悬菌液在不同真空压力条件下侵染5 min、10 min、15 min、20 min、30 min,侵染结束后将愈伤组织置于带滤纸的培养皿中,吸去多余菌液后放在共培养基上,21℃共培养3 d。
3.2.3转化植株的获得
3.2.3.1抗性愈伤组织的的筛选
共培养3 d后的愈伤组织转移至恢复培养基中,恢复培养4 d后转移至筛选培养基中筛选抗性愈伤组织。每14 d筛选一次,3次筛选后得到抗性愈伤组织。
3.2.3.2抗性愈伤组织的分化
筛选得到的抗性愈伤组织置于分化培养基中,在27℃,2000Lx光强,每天光照16 h的条件下分化生长,直至得到幼苗。
3.2.3.3 壮苗培养和移栽
将幼苗放在生根培养基中生长,待根系发达后,敞开瓶口炼苗,1周后移栽于温室。
3.2.3 转化植株的PCR检测
3.2.3.1 DNA的提取
当转化植株处于4叶期时取玉米幼嫩的叶片,用CTAB法提取转基因玉米的基因组DNA。
3.2.3.2 PCR检测抗性植株
根据HAL1的基因序列设计引物PCR 引物: PR :5’CCAGAAACCCACGTCATGCC3’ PF:5’CAGGAACCGCAGGAGTGGA3’;反应体系为20 μl。PCR扩增序为:(1) 95℃,预变性 5 min;(2) 95℃,变性1 min;(3) 56℃,退火 30s;(4) 72℃,延伸 30s;(5)重复步骤(2)~(4)40次;(6)72℃延伸10 min。PCR产物大约375bp。
作者贡献
孙传波是本研究的实验设计和论文初稿的写作,孙传波、郭嘉、陶蕊、孟凡梅和曲文利参与实验研究的执行和数据分析;韦正乙提供植物表达载体,李海华负责转化植株在责温室的生长,刘文国和袁英是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由国家转基因生物新品种培育重大专项课题(2009ZX08003-018B)资助。
参考文献
Bordas M., Montesinos C., Dabauza M., Salvador A., Roig LA., Serrano R., and Moreno V., 1997, Transfer of the yeast salt tolerance gene HAL1 to Cucumis melo L. cultivars and in vitro evaluation of salt tolerance, Transgenic Res., 6: 41-50 doi:10.1023/A:1018453032336 PMid:9032977
Gould J., Devey M., Hasegawa O., Ulian E.C., Peterson G., and Smith R.H., 1991, Transformation of Zea mays L. using Agrobacterium tuniefaciens and the shoot apex, Plant Physiol., 95: 426-434 doi:10.1104/pp.95.2.426 PMid:16668001 PMCid:1077548
Huang L., and Wei Z.M., 1999, Agrobacterium tumefaciens mediated maize transformation, Shiyan Shengwu Xuebao (Acta Biologiae Experimentalis Sinica), 32(4): 381-387 (黄璐, 卫志明, 农杆菌介导的玉米遗传转化, 实验生物学报, 1999, 32(4): 381-387)
Huang S., Gilbertson L.A., Adams T.H., Malloy K.P., Reisenbigler E.K., Birr D.H., Snyder M.W., Zhang Q., and Luethy M.H., 2004, Generation of marker free transgenic maize by regular two2border Agrobacterium transformation vectors, Transgenic Res., 13(5): 451-461 doi:10.1007/s11248-004-1453-3 PMid:15587269
Ishida Y., Saito H., Ohta S., Hiei Y., Komari T., and Kumashiro T., 1996, High efficiency transformation of Zea mays L. mediated by Agrobacterium tumefaciens, Nature Biotechnology, 14(6): 745-750 doi:10.1038/nbt0696-745 PMid:9630983
Rhodes C.A., Pierce D.A., Mettler I.J., Mascarenhs D., and Detmer J.J., 1988, Genetically transformed maize plants from protoplasts, Science, 240: 204-207 doi:10.1126/science.2832947 PMid:2832947
Rios G., Ferrando A., and Serrano R., 1997, Mechanisms of salt tolerance conferred by over expression of the HAL1 gene in Saccharomyces cerevisiae, Yeast, 13: 515-528 doi:10.1002/(SICI)1097-0061(199705)13:6<515::AID-YEA102>3.3.CO;2-O
Rohila J.S., Jain G.K., and Wu R., 2002, Genetic improvement of Basmati rice for salt and drought tolerance by regulated expression of a barley HAL1 cDNA, Plant Sci., 163(3): 525-532
doi:10.1016/S0168-9452(02)00155-3
Tian J.L., Yang Y.A., He Y.K., 2003, Salt tolerance of Transgenic Tomato with HAL1 Gene, Zhiwu Shengli Yu Fenzi Shengwuxuebao (Journal of Plant Physiology and Molecular Biology), 29(5): 409-414(田吉林, 杨玉爱, 何玉科, 2003, 转HAL1基因番茄的耐盐性, 植物生理与分子生物学学报, 29(5): 409-414)
Yang S.X., Zhao Y.X., Zhang Q., He Y.K., Zhang H., and Luo D., 2001, HAL1 mediate salt adaptation in Arabidopsis thaliana, Cell Res, 11 (2): 142-148 doi:10.1038/sj.cr.7290079 PMid:11453546
Yuan Y., Li Q.Y., Hao W.Y., Tan H., Kong X.M., Zhang G.D., and Liu D.P., 2006, Studies on Influencing Factors of Agrobacterium tumefaciens mediated Maize Transformation, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 4(2): 228-232 (袁英, 李启云, 郝文媛, 谭化, 孔祥梅, 张光弟, 刘德璞, 2006, 农杆菌介导的玉米遗传转化影响因子的研究, 分子植物育种, 4(2): 228-232)